|
|
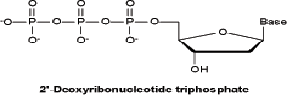
DNA replication A. Semiconservative replication 1. As shown by Meselson and Stahl in 1958, when DNA is replicated, each daughter molecule consists of one old strand and one new strand. This is the so-called semiconservative model. MESELSON, MATTHEW, & FRANKLIN W. STAHL. The replication of DNA in Escherichia coli. Proceedings of the National Academy of Sciences USA 44(7): 671–682, 1958. B. The immediate precursors of DNA are the four deoxynucleoside triphosphates, dATP, dGTP, dCTP, and dTTP.
1. The four deoxynucleotide triphosphates can be made from central metabolites in Bacteria, Archaea, and most eukaryotes. 2. For all four compounds, the 2’ oxygen is removed from the corresponding ribonucleotide diphosphate (ADP, GDP, CDP, and UDP) by an enzyme called ribonucleotide reductase, which produces dADP, dGDP, dCDP, and dUDP as products. a) Hydroxyurea inhibits this reaction and is occasionally used as an antimicrobial agent, but it’s not terribly specific. 3. Three out of the four (dADP, dGDP, and dCDP) are then phospho- rylated by a kinase to produce the final dATP, dGTP, and dCTP. But poor dUDP needs a methyl group in the 5 position. 4. Oddly enough, in yet another example of inefficiency in biological design, dUDP is subjected to the action of a phosphatase to convert it to dUMP, then methylated by thymidylate synthetase to dTMP, then subjected to the action of a kinase (twice) to produce dTTP. a) Thymidylate synthetase is inhibited by 5-fluorodeoxyuridine, which has been used as an antimicrobial agent. Again, it has specificity problems. b) Thymidylate synthetase gets the methyl group from the coenzyme tetrahydrofolate. (1) The biosynthesis of dTMP, and hence DNA, is apparently the single most significant drain on tetrahydrofolate in thecell. (2) Tetrahydrofolate is regenerated by an enzyme called dihy-drofolate reductase, which can be inhibited by trimetho-prim, used as an antimicrobial compound, and metho-trexate, used as a cancer chemotherapeutic agent, a treatment for rheumatoid arthritis, and an abortifactant.
C. Deoxyribonucleotide polymerization
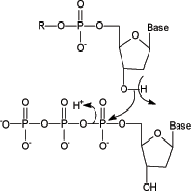
1. The actual polymerization reaction is a simple nucleophilic attack of the free 3’-hydroxyl group on the -phosphorus of the incoming deoxynucleotide triphosphate, displacing pyrophosphate. This was postulated by Kornberg in 1958. (BESSMAN, MAURICE J., I. R. LEHMAN, ERNEST S. SIMMS, & ARTHUR KORNBERG. Enzymatic synthesis of deoxyribonucleic acid II. General properties of the reaction. Journal of Biological Chemistry 233(1):171–177, July 1958.) 2. The replication system requires primers to add onto; most (not all) DNA polymerases cannot start a chain de novo. When necessary, RNA primers are used. 3. The replication system requires an antiparallel, complementary template strand to determine the nucleotide sequence of the new strand. 4. And, of course, the replication requires the four deoxynucleotide triphosphates: dATP, dGTP, dCTP, and dTTP.
Table 1-1. Proteins involved in E. coli DNA replication
D. Replication of double-stranded DNA 1. The replication system is inordinately complex (see overhead “Some Components of the Replication Fork” and Table 1-1). 2. Leading strand. 3. Lagging strand a) Okazaki fragments: 10–12-nucleotide RNA primers with DNA added on b) 5’ exonuclease activity of DNA polymerase I removes RNA, and polymerase activity fills in DNA c) Ligase ties everything together E. Replication errors are a source of mutations, but the replication error rate is low 1. Editing a) PolA (DNA polymerase I) b) DnaQ 2. Methyl-directed mismatch repair a) Mismatches can be repaired with an efficiency greater than 50% because the mismatch repair system can tell the old strand from the new strand: the old strand is more highly methylated at the 6 position of the adenines in palindromic GATC/CTAG sequences 3. Role of editing and mismatch repair in maintaining replication fidelity a) The replication error rate for wild-type E. coli appears to be about 4.7 x 10 -4 substitutions per genome per cell division, or one error in every 2,000 progeny. F. Relationship of DNA replication and cell division 1. Structure of the bacterial chromosome a) The bacterial cell is genetically haploid, even if it has more than one copy of the chromosome per cell. b) There is only one allele for every locus. c) Most bacterial chromosomes are covalently closed loops, or circles, of double-stranded (1) Borrelia burgdorferi, the causative agent of Lyme disease, has a linear chromosome (and linear plasmids). 2. Replication of the bacterial chromosome a) Initiation takes place at oriC, the origin of chromosomal replication, and proceeds in the two opposite directions with two complete replication systems.(CAIRNS, JOHN. The bacterial chromosome and its manner of replication as seen by autoradiography.Journal of Molecular Biology 6: 208–213, 1963.) b) Termination occurs when the two replication forks meet somewhere near the terA terB region c) Two daughter molecules need to be resolved (untangled), then partitioned 3. Coordination of DNA replication with the cell cycle a) It appears that replication of the E. coli chromosome takes 40 min. b) Under favorable conditions of growth, a new round of replication is initiated before the old round is terminated. Two or three pairs of replication forks may be moving at any given instant. c) Thus E. coli can have a generation time smaller than its DNA replication time! G. Antibiotics that modify DNA structure and/or inhibit replication 1. Drugs that block precursor biosynthesis a) Trimethoprim and methotrexate inhibit dihydrofolate reductase. b) Hydroxyurea inhibits ribonucleotide reductase c) 5-Fluorodeoxyuridine and 5-fluorouridine compete with the natural substrate, dUMP, for thymidylate synthetase and thus inhibit its action. 2. Antibiotics that block polymerization of nucleotides a) Dideoxynucleotides have no 3’ hydroxyl group, and thus act as chain terminators. They don’t make very useful antibacterial agents, but they have other technological uses, e.g. as antiviral agents and in DNA sequencing. 3. Antibiotics that affect DNA structure a) Mitomycin C (made by Streptomyces caespitosus) cross-links nearby guanine bases, thus disrupting replication. b) Acridine dyes such as proflavin, ethidium, and chloroquin intercalate between base pairs and cause deletions and insertions, resulting in frameshift mutations. c) 5-Bromouracil will substitute for thymine in DNA synthesis, but has a tendency to basepair a bit more like cytosine than thymine, resulting in substitution mutations. It also makes DNA more susceptible to ultraviolet light damage. 4. Antibiotics that inhibit DNA gyrase a) Nalidixic acid, oxolinic acid, chloromycetin, and ciprofloxacin all inhibit the action of GyrA. b) Novobiocin and coumermycin inhibit the action of GyrB. III. Does the bacterial genome really consist of a single, circular molecule of DNA, the BIO 101 model? A. Certainly 99% of the genes of Escherichia coli map to a single, circular molecule of DNA 1. There is physical evidence for a circular chromosome 2. There is genetic evidence for a circular chromosome (WOLLMAN, E. L., F. JACOB, & W. HAYES. Conjugation and genetic recombination in Escherichia coli K-12.Cold Spring Harbor Symposia on Quantitative Biology 21: 141–162, 1956) B. However, now that several genome projects have come to conclusion, the picture is by no means uniform… Table 1-2. Replicons in some sequenced bacterial genomes with multiple heritable elements
|
|
BIOTECH PORTAL |